Touch and the PIEZO ion channels - their role in pain

When a manual touch therapist places their hands on the skin of a client, a sophisticated chain of biological events begins. What may feel like heat, pressure, movement or stretching is detected by specialised tissue receptors located in and beneath the skin. Among these are mechanosensitive ion channels known as PIEZO1 and PIEZO2, and thermochemical sensing channels called TRPV. These channels were only discovered in the past two decades, and their significance in sensory detection was so impactful that Ardem Patapoutian and David Julius were awarded the Nobel Prize for Physiology or Medicine in 2021 for this work (The Nobel Prize, 2021). A 2025 article in Nature discusses how these channels adapt and influence the experience of pain.
PIEZO and TRPV channels detect vital changes in the environment but respond to different types of stimuli. PIEZO channels are mechanosensitive, meaning they detect pressure, stretch, touch and proprioception (Zhao et al., 2018). TRPV channels, particularly TRPV1, are thermosensitive and chemosensitive, detecting heat, chemical irritants and inflammation (Dubin and Patapoutian, 2010). Although both channel types share the purpose of protecting the body and maintaining homeostasis, their biological mechanisms and roles are distinct. This distinction also challenges and refines aspects of the traditional Gate Control Theory of Pain from Melzack in Wall in 1965. Originally, the theory proposed that non-noxious input from mechanoreceptors could inhibit nociceptive input at the spinal cord, effectively 'closing the gate' to pain. Regarded as being over simplistic in current pain science, the Gate Control idea of closing the gate to pain prevails. However, further research suggests that PIEZO and TRPV channels may operate in a more complex and context-dependent manner expanding how nociception is processed. For example, under inflammatory conditions, PIEZO2 can become involved in nociceptive signalling, meaning that the same input once perceived as non-threatening could now be processed as damaging resulting in pain. Rather than simply opening or closing a gate, these channels illustrate how input is dynamically interpreted depending on the physiological state of the tissue and central nervous system, commonly called central sensitisation.
|
Feature |
Piezo1 and Piezo2 |
TRPV Channels (especially TRPV1) |
|
Main Stimulus |
Mechanical pressure, stretch |
Heat, chemical irritation, some mechanical |
|
Primary Role |
Touch, proprioception, tissue homeostasis |
Noxious stimuli and inflammation signalling |
|
Key Locations |
Skin, blood vessels, connective tissues, sensory neurones |
Sensory neurones, skin, mucosal tissues |
|
Pain Involvement |
Mechanical allodynia (inflammation linked) |
Pain because of thermal and chemical change and inflammation |
|
Channel Structure |
Trimeric, dome shaped |
Tetrameric, smaller and flexible |
PIEZO channels are embedded in the membranes of various cells. Structurally, they are large, trimeric complexes shaped like three-bladed propellers. When mechanical force is applied, the blades shift apart, opening the central pore and allowing positively charged ions such as calcium (Ca²⁺) and sodium (Na⁺) to enter the cell (Zhao et al., 2018).
PIEZO1 and PIEZO2 serve different roles. In non-neuronal tissues, PIEZO1 channels in the skin influence barrier function and healing. In blood vessels, they regulate vascular tone. In fibroblasts, they impact tissue repair and matrix production (Botello-Smith et al., 2019; Douguet et al., 2019; He et al., 2023; Murthy et al., 2017).
PIEZO2 is predominantly found in sensory neurones, particularly mechanosensitive types such as Aβ low-threshold mechanoreceptors (LTMRs), Aδ-LTMRs, and proprioceptive neurones located in muscle spindles and Golgi tendon organs. These neurones are responsible for detecting light touch, vibration and proprioception. PIEZO2 is essential for translating subtle mechanical stimuli into electrical signals interpreted by the nervous system (Szczot et al., 2018, 2021). When activated, these channels generate an action potential, an electrical signal that travels along the neurone, carrying information to the spinal cord’s dorsal horn. Here, neurotransmitters are released, relaying the signal through the central nervous system to the brain, where it is consciously perceived.
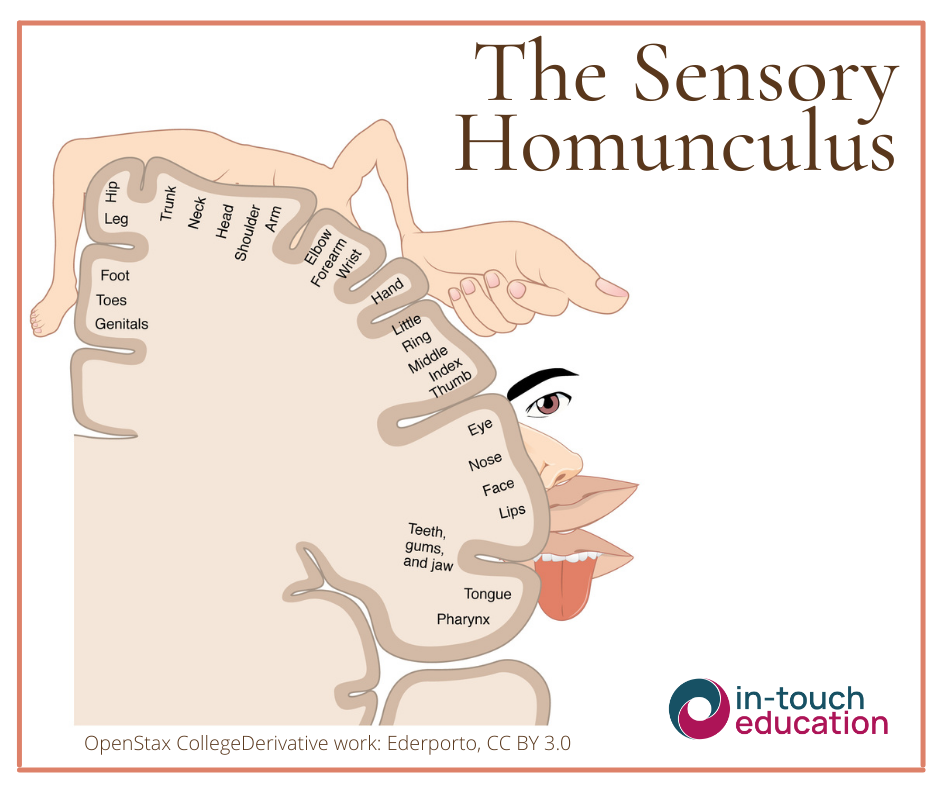
This sensory information is processed through ascending tracts to the brainstem, thalamus, and ultimately the primary somatosensory cortex in the parietal lobe. The brain creates a detailed sensory map, known as the homunculus, with areas such as the hands and face receiving the largest representation (Boldrey and Penfield, 1937). The perception of touch is shaped not only by its physical properties but by memory, emotion and context.
Merkel cells, found in both hairy and hairless skin, form complexes with Aβ-LTMRs. This Merkel-neurite complex is key for detecting fine textures and sustained pressure (Woo et al., 2015). Both Merkel cells and Aβ neurones contain PIEZO2. The Merkel cell responds to continuous pressure, while the neurone detects movement. This system underpins slowly adapting responses and highlights PIEZO2’s crucial role in discriminative touch (Szczot et al., 2021).
Nociceptors are specialised sensory neurones distinct from mechanoreceptors. While mechanoreceptors, such as Aβ and Aδ-LTMRs, respond to non-threatening mechanical input like light touch, vibration and proprioception, nociceptors are designed to detect potentially harmful or noxious stimuli. These include thermal, mechanical or chemical threats that may signal tissue damage. Nociceptors are primarily associated with Aδ and C fibres and are responsible for transmitting nociceptive input to the central nervous system, not pain itself but the raw signals that may result in the brain constructing a pain experience.
During inflammation, PIEZO2 channels can become involved in this nociceptive processing. Prostaglandin E2 (PGE2), an inflammatory mediator, sensitises tissues and can cause PIEZO2-expressing neurones to fire spontaneously. This spontaneous activity, particularly in Aδ and C fibres, sends ongoing nociceptive input to the central nervous system, even without external stimuli (Ghitani et al., 2025). When normal mechanosensory LTMR signals from touch arrive at the spinal cord alongside this background nociceptive input, the sensory system may misinterpret the signal as a potential threat. This misprocessing contributes to mechanical allodynia, a condition in which non-noxious stimuli, such as gentle touch or brushing, are perceived as threatening or unpleasant. Unlike normal protective responses to noxious stimuli, mechanical allodynia involves the brain misinterpreting innocuous input due to altered central processing. This can significantly affect quality of life, as everyday physical contact may provoke distress or discomfort. It is a hallmark of central sensitisation and often persists in chronic and persistent pain states, posing a challenge for both clients and clinicians in managing sensory experiences that no longer match the original input.
Importantly, LTMR responses remain unchanged during inflammation. It is the nociceptor’s spontaneous activity that alters the interpretation of touch at the central level. If PIEZO2 is genetically removed or silenced (called knockout), mechanical allodynia does not occur, suggesting PIEZO2 activity in Merkel cells and mechanoreceptors is critical in touch-evoked sensory experiences (Ghitani et al., 2025).

PIEZO2 channels appear to serve as key modulators in the interaction between mechanical input and nociceptive processing, particularly in inflammatory states. Manual therapists, by applying skilled and intentional mechanical force, may be engaging with these very channels. In non-inflamed tissues, activating PIEZO1 and PIEZO2 may support blood flow, proprioception and tissue responsiveness. In sensitised systems, where PIEZO2 contributes to altered central interpretation of input, there is theoretical potential for manual therapy to modulate that input. Therapeutic touch could, depending on context and method, influence both peripheral activation of PIEZO channels and central integration of those signals. This reframes manual therapy as a sensory intervention capable of modulating neurobiological communication, potentially supporting adaptation and restoration.
It is not about a specific technique, branded approach or protocol that determines therapeutic value, as all touch generates the same type of physiological response, depending on context. What matters more is the way in which a technique is delivered, how the touch is applied and the context in which it occurs. These factors can significantly influence the quality and outcome of the therapeutic interaction. Further research is essential, but emerging evidence positions skilled touch as a valid route for engaging with the sensory systems that contribute to both homeostasis and nociceptive modulation.
References
Boldrey, E.P.W., 1937. Somatic Motor and Sensory Representation in the Cerebral Cortex of Man as Studied by Electrical Stimulation.
Botello-Smith, W.M., Jiang, W., Zhang, H., Ozkan, A.D., Lin, Y.C., Pham, C.N., Lacroix, J.J., Luo, Y., 2019. A mechanism for the activation of the mechanosensitive Piezo1 channel by the small molecule Yoda1. Nat Commun 10. https://doi.org/10.1038/S41467-019-12501-1
Coste, B., Mathur, J., Schmidt, M., Earley, T.J., Ranade, S., Petrus, M.J., Dubin, A.E., Patapoutian, A., 2010. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science (1979) 330, 55–60. https://doi.org/10.1126/science.1193270
Douguet, D., Patel, A., Xu, A., Vanhoutte, P.M., Honoré, E., 2019. Piezo Ion Channels in Cardiovascular Mechanobiology. Trends Pharmacol Sci 40, 956–970. https://doi.org/10.1016/J.TIPS.2019.10.002
Dubin, A.E., Patapoutian, A., 2010. Nociceptors: the sensors of the pain pathway. J Clin Invest 120, 3760. https://doi.org/10.1172/JCI42843
Ghitani, N., von Buchholtz, L.J., MacDonald, D.I., Falgairolle, M., Nguyen, M.Q., Licholai, J.A., Ryba, N.J.P., Chesler, A.T., 2025. A distributed coding logic for thermosensation and inflammatory pain. Nature 2025 1–8. https://doi.org/10.1038/s41586-025-08875-6
He, J., Fang, B., Shan, S., Li, Q., 2023. Mechanical stiffness promotes skin fibrosis through Piezo1-mediated arginine and proline metabolism. Cell Death Discov 9, 1–9. https://doi.org/10.1038/S41420-023-01656-Y;SUBJMETA=1642,304,319,443,631,80;KWRD=MECHANISMS+OF+DISEASE,METABOLIC+DISEASES
Lei, L., Wen, Z., Cao, M., Zhang, H., Ling, S.K.K., Fu, B.S.C., Qin, L., Xu, J., Yung, P.S.H., 2024. The emerging role of Piezo1 in the musculoskeletal system and disease. Theranostics 14, 3963–3983. https://doi.org/10.7150/THNO.96959
Murthy, S.E., Dubin, A.E., Patapoutian, A., 2017. Piezos thrive under pressure: Mechanically activated ion channels in health and disease. Nat Rev Mol Cell Biol 18, 771–783. https://doi.org/10.1038/nrm.2017.92
Savadipour, A., Palmer, D., Ely, E. V., Collins, K.H., Garcia-Castorena, J.M., Harissa, Z., Kim, Y.S., Oestrich, A., Qu, F., Rashidi, N., Guilak, F., 2023. The role of PIEZO ion channels in the musculoskeletal system. Am J Physiol Cell Physiol 324, C728. https://doi.org/10.1152/AJPCELL.00544.2022
Szczot, M., Liljencrantz, J., Ghitani, N., Barik, A., Lam, R., Thompson, J.H., Bharucha-Goebel, D., Saade, D., Necaise, A., Donkervoort, S., Foley, A.R., Gordon, T., Case, L., Bushnell, M.C., Bönnemann, C.G., Chesler, A.T., 2018. PIEZO2 mediates injury-induced tactile pain in mice and humans, Sci. Transl. Med.
Szczot, M., Nickolls, A.R., Lam, R.M., Chesler, A.T., 2021. The Form and Function of PIEZO2. Annu Rev Biochem 90, 507. https://doi.org/10.1146/ANNUREV-BIOCHEM-081720-023244
Woo, S.H., Lumpkin, E.A., Patapoutian, A., 2015. Merkel cells and neurons keep in touch. Trends Cell Biol. https://doi.org/10.1016/j.tcb.2014.10.003
Zhao, Q., Zhou, H., Chi, S., Wang, Y., Wang, Jianhua, Geng, J., Wu, K., Liu, W., Zhang, T., Dong, M.Q., Wang, Jiawei, Li, X., Xiao, B., 2018. Structure and mechanogating mechanism of the Piezo1 channel. Nature 554, 487–492. https://doi.org/10.1038/nature25743
